
It's tempting, and easy, to think of our bodies as constant and that growth is only occuring when we can see it. However, the body is continually tearing itself down and building itself back up. The entire intestinal epithelium regenerates completely, a couple cells at a time, over the course of 4 days to 1 week. Our skin is continually growing outwards and being worn away. Some organ systems, such as skeletal bone, rebuild themselves at a much slower rate while others, such as the central nervous system, are essentially immutable over the entire course of our lives. Most of the molecules that make up the body are in a continual state of flux as metabolism breaks down cellular products in adipocytes and delivers free fatty acids to the liver for oxidation into pyruvate, phosphocreatinine, or glycogen for use by skeletal muscle to do work, although this is not the only direction or only pathway in which such conversions constantly take place.
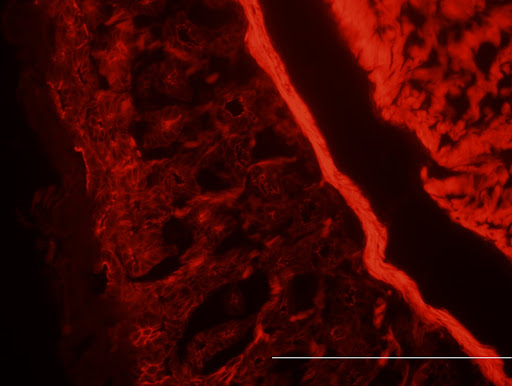 Colonic smooth muscle actin (red). The bright red strand is the smooth muscle lining the columnar epithelium of the colon (fainter chambered red). The really bright red strand is the musclaris mucosa that helps to drive peristalsis, or the sequential movement of intestinal smooth muscles to drive lumenal contents (food being digested) along the length of the gastrointestinal tract. 40X magnification, bar is 200um.
Colonic smooth muscle actin (red). The bright red strand is the smooth muscle lining the columnar epithelium of the colon (fainter chambered red). The really bright red strand is the musclaris mucosa that helps to drive peristalsis, or the sequential movement of intestinal smooth muscles to drive lumenal contents (food being digested) along the length of the gastrointestinal tract. 40X magnification, bar is 200um.Muscles are made up of highly-organized fibers, such as the actin in the picture above. These fibers are organized into bundles that are studded with molecular motors known as kinesins that effectively run along parallel fibers. When you contract a muscle and tense it up, the fibers are dragged past each other by the kinesins to make the effective length of the muscle shorter and producing work in the form of moving whatever the muscle is attached to. In the process of this, the molecular motors burn cellular energy in the form of adenosine triphosphate (ATP) that the body makes from pyruvate molecules.
Over time as the muscle fibers get used they start to breakdown due to mechanical wear and tear (the initial burn from a workout is due to accumulation of calcium from anaerobic respiration; the soreness the next day is due to injured muscle fibers). The order of the molecules in the fibers starts to break down, and it's much cheaper in terms of energy used for the body to break down the worn-out muscle fibers and replace them with new fibers than it is to just repair them. As such, skeletal muscles are in a continual flux of breakdown and new fiber synthesis. This dynamic process that continually renews our muscles is not only completely normal, it's advantageous. Genetic experiments with mice have shown that deleting the gene that encodes myostatin, a protein strongly implicated in muscle breakdown, results in massively muscled mice who aren't any stronger than their scrawny companions that still have myostatin. Deletion of myostatin arrests muscle breakdown, but also leads to hypertrophy of those muscles primarily through retention of damaged and useless muscle fibers that add absolutely nothing to the ability of the muscles to perform work (strength).
In relation to food, the breakdown and building of muscles, which are really just facets of the catabolic and anabolic states of metabolism, can be classified more simply as muscle protein breakdown (MPB) and muscle protein synthesis (MPS). Put simply, muscles will shrink when the rate of MPB exceeds that of MPS and vice versa. The body increases MPB when blood sugar and insulin levels are low and the bodily stores of glycogen in liver and skeletal muscle have been depleted. Although fatty acids are also mobilized for energy from fat tissues, the net effect of this is that the body begins to slowly eat at its skeletal muscles to keep going. The body also pulls acidic or basic amino acids out of muscle proteins as needed to help regulate blood pH, which has to remain within a narrow range for us to survive. After eating, and for 4-6h afterwards, the body decreases MPB and increases MPS such that new muscle fibers are built. How we work out, the spacing of our meals, and even the composition of those meals all influence the efficiency with which muscle protein synthesis occurs. For what it's worth, branched amino acids such as valine, leucine, and isoleucine are absorbed across the brush border of the duodenum and into the blood stream at a faster rate than other free amino acids.
The primary lesson of all of this is that bodily flux is entirely normal. I know that when I first started trying to gain weight I obsessively checked the scale everyday and would be disappointed when it registered a 2kg loss over the course of a day, and astounded when it would suddenly report a 4kg increase in 1 day. The reality is that one's immediate weight, and muscle mass, are determined by a large number of factors and unless we are trying to be super-athletes we shouldn't sweat them too much. Variation is normal. The body already has flux down and we don't even have to think about it for it to occur.
P.S. - Scicurious of Neurotopia has published an excellent introduction to the mechanics of neurotransmission, which are quite relevant to how nerve signals are transduced to musclar action through neuromusclar junctions.
[Sources for this post included what I remember from a few old textbooks (primarily Griffin and Ojedas' "Textbook of Endocrine Physiology", Vander, Sherman, and Lucianos' "Human Physiology", maybe some Alberts et als' "Molecular Biology of the Cell", and a wide scattering of abstracts I remember reading, but not saving.]




1 comment:
Srsly Toaster, if you post another beautiful picture with fracking scale bar sliding off the side of the image Ima gonna scream!
Post a Comment