
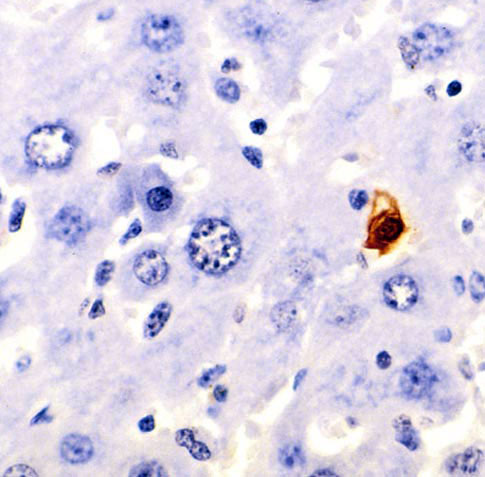
 Figure A: TUNEL histochemical staining in murine liver, brown cell is apoptotic.
Figure A: TUNEL histochemical staining in murine liver, brown cell is apoptotic.The apoptotic program ultimately results in highly oxidative and degradative enzymes (such as proteases) hidden away in the mitochondria being released into the cytoplasm to wreak havoc. Usually the raw material of a dying cell is tidily absorbed by its neighbors to be recycled. I've always imagined mitochondria as pulsing with a low, gentle buzz in normal cellular physiology with occassional metallic pings as statistical flucuations in the net free energy of electrons falling down the electron transport chain is captured in ATP. Following this, I think the sound of caspase-8 et al slicing open the mitochondria would be like the initial panicked braking shriek of a train loaded with Furbies who are quickly drowned out in the self-amplifying roar like a tornado grinding through a gravel pit as the apoptotic effector enzymes set to work dissolving the cell from within.
Apoptosis is absolutely essential not just to adult homeostasis, but also to normal ontogeny. Without apoptosis organs would fail to separate, fingers would remain stuck together, and many other things would go very, very wrong. There are 2 families of intracellular proteins that battle to determine whether or not a cell will become apoptotic: the (generally) pro-apoptotic Bcl-2 family and the (generally) anti-apoptotic IAP family. Conveniently, IAP stands for Inhibitor of Apoptosis Protein. A recent review by Dr. O'Riordan et al discussed the diverse and essential roles for IAP proteins in normal tissue development across a wide range of model organisms. From ablated organ development in the absence of Diap1 in Drosophila larvae to stunted hematopoeitic developmental repertoire in the abscence of Survivin in mice, IAPs seem to be evolutionarily conserved signal transducers that integrate diverse extracellular signals into a coherent cellular action. Developmentally, the IAP proteins seem to be involved in everything from proper vascularization to chromosome stability, although it is important to note that direct modulation of apoptosis in developmental processes has only been established in invertebrates. Lack of any one of several IAPs in higher chordates has not been directly linked to developmental apoptosis, but several abnormal embryonic phenotypes and attenuated adult functional capacities have been demonstrated.
IAPs are grouped by the prescence of BIRs (baculovirus IAP repeats) and many also have RING domains. Both motifs have been found to have zinc-finger conformations and the interaction of different sections of adjacent BIR motifs in some proteins, such as direct inhibition of pro-apoptotic caspases-3 and -7 by BIR2 of XIAP (X-linked inhibitor of apoptosis protein), has been found to modulate a number of diverse effects. These diverse effects are potentiated by the ubiquitin ligase activity that some RING domains have demonstrated. IAPs help the organism balance necessary apoptosis and unnecessary apoptosis, and because apoptosis is required for the homeostasis of most tissues the IAP family has been evolutionarily conserved and biochemically diversified. IAPs remain an active and engaging area of research that holds great promise in the treatment of pathologies from cancer to intracellular bacterial infections and underscore how a little sacrifice for the team by one cell can make a massive impact on the constituent organism's overall fitness.
IAPs have also been found to modulate innate immunity, which will be discussed in another post.
ORIORDAN, M., BAULER, L., SCOTT, F., & DUCKETT, C. (2008). Inhibitor of Apoptosis Proteins in Eukaryotic Evolution and Development: A Model of Thematic Conservation Developmental Cell, 15 (4), 497-508 DOI: 10.1016/j.devcel.2008.09.012
Additional Source: Molecular Biology of the Cell; Alberts et al; 4th ed.; pages 1010-1014



